Птерозавры: история крылатых ящеров
Размах крыльев
Каких размеров может достигать животное, освоившее полет? Казалось бы, ответ у нас перед глазами, достаточно взглянуть на крупнейших из современных летающих птиц. Размах крыльев альбатросов и андских кондоров превышает 3,5 метра. При таких размерах у этих птиц удивительно небольшой вес - 11 кг у альбатроса и 15 у кондора. Лебеди, чей размах крыльев может превышать 2,4 м, могут иметь и 20 кг, но такой вес заметно сказывается на их летных способностях.
И все же, история жизни на Земле показывает, что это не предел. Пелагорнис (Pelagornis sandersi), живший 25 млн лет назад на восточном побережье Северной Америки, достигал 7,4 м в размахе крыльев. С ним соперничает аргентавис (Argentavis magnificens) из позднего миоцена Аргентины, имевший сходные размеры. Однако, и эти птицы уступают первенство в размерах представителям вымершей группы рептилий - птерозаврам.

Окаменелые остатки летающих ящеров были описаны еще в конце XVIII века. Итальянский натуралист Козимо Коллини посчитал, что несуразный скелет с длинными лапами принадлежит водному животному. Подобные представления бытовали даже спустя 40 лет, но большинство палеонтологов к тому времени рассматривали птерозавров как аналогов летучих мышей. Находки были редкими и фрагментарными, что затрудняло изучение этих животных. Мелкие, теплокровные, покрытые шерстью то-ли ящеры, то-ли предки птиц - так изображали птерозавров до 70-х годов XIX века.
Ситуацию изменили окаменелости из Америки. Тысячи скелетов и их фрагментов, хорошая сохранность, и - огромные размеры. Отниел Чарльз Марш в меловых отложениях Канзаса нашел семиметрового птеранодона. И это была не единственная находка. Постепенно маятник представлений о птерозаврах сместился (как и в случае с динозаврами) в “рептильную” сторону. Их стали рассматривать как курьезы природы, неудавшиеся эксперименты эволюции. Холоднокровные, чешуйчатые, медлительные, способные только парить в небе, спрыгивая с прибрежных скал, они были незаслуженно забыты до 1960-х годов.
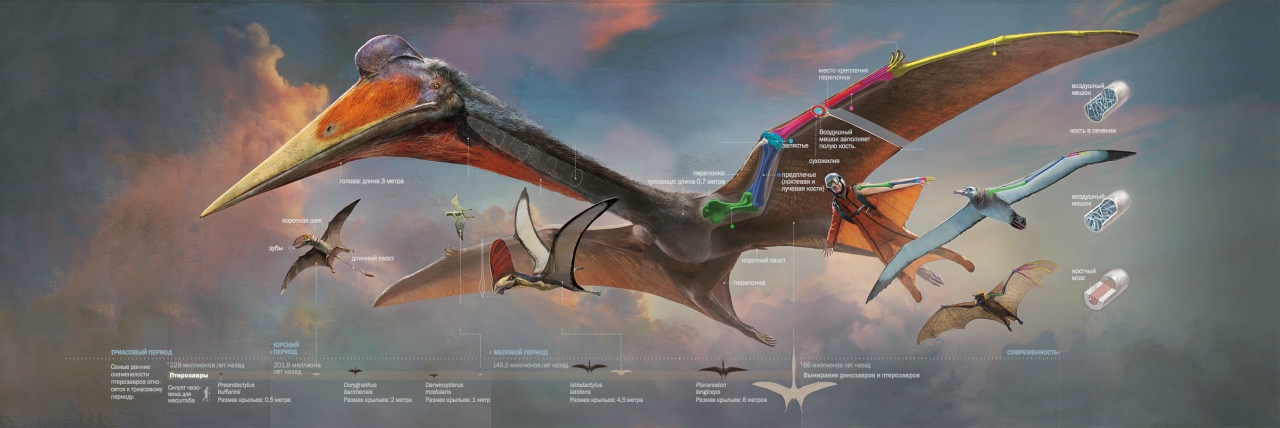
Возрождение интереса к динозаврам, начавшееся с исследований Джона Острома, захватило и их летающих соседей по мезозою. Новые находки в Бразилии и Китае, а также новые методы изучения позволили пересмотреть представления о птерозаврах. А в 1975 году открытие кетцалькоатля стало началом изучения неизвестного ранее семейства аждархид. Его представители с размахом крыльев до 12 м (а может, и более) - крупнейшие животные, когда либо поднимавшиеся в воздух.
Или не поднимавшиеся? Механика полета гигантских птерозавров, равно как и оценка их массы и размеров, были предметом споров со времен их открытия. Сложность состоит в том, что сейчас нет животных, которых можно было бы использовать в качестве аналогов птерозавров. Ни птицы, ни тем более летучие мыши таковыми не являются, слишком велика разница в анатомии. И все же, за неимением лучшего, на протяжении долгого времени именно птицы использовались для моделирования биомеханики птерозавров.
Выводы на основе такой замены оказались не утешительными. Весовой рекорд среди летающих птиц принадлежит дрофам, и составляет 22 кг. Птерозавр такого веса должен был иметь размах крыльев 4,2 метра. При большем размахе крыльев он мог бы подняться в воздух, только имея экстремально низкий вес, около 16 кг для 7 м и 70 кг для 10 м. При этом непонятно, как много мускулатуры останется при таком снижении веса. Критическое соотношение массы и размаха крыльев оценивается некоторыми исследователями в 41 кг и 5,1 м. Выше этого предела животное становится слишком тяжелым для полета.
Эти расчеты входят в явное противоречие с существованием птеранодонов, орнитохейрусов и аждархид, чьи размеры превышают указанный предел. Один из крупнейших видов, кетцалькоатль (Quetzalcoatlus northropi), по современным оценкам имел размах крыльев до 10 - 12 м и весил 200 - 250 кг. Строение тела гигантских птерозавров не оставляет сомнений в их способности летать. Ключ к ответу на вопрос, как птерозаврам удалось стать крупнейшими летающими животными, кроется в их специфической анатомии. А понимание анатомии невозможно без знакомства с эволюционной историей.

Драконы учатся летать
Истоки эволюции птерозавров теряются в туманных лесах триаса. Проблема здесь, в общем, та же, что и с летучими мышами. Самые древние известные птерозавры уже максимально приспособлены для полета. А те животные, которые предлагаются на роль возможных предков, летными характеристиками не обладают.
Тем не менее, установлено, что птерозавры с большой долей вероятности являются архозавроморфами, то есть, приходятся родней динозаврам, а значит, и птицам. Правда, родство это не близкое. Предки птерозавров пошли своим эволюционным путем, отделившись от динозавроподобных рептилий, еще до того, как последние стали собственно динозаврами.
Кого только не предлагали на роль прародителей птерозавров! Среди них отметились даже танистрофеиды, длинношеие водоплавающие архозавроморфы.
Одним из кандидатов был шаровиптерикс (Sharovipteryx mirabilis). Это мелкая, длиной 20 см, рептилия из Ферганской долины, освоившая парящий полет. Правда, летная перепонка шаровиптерикса располагалась между задними лапами, а передние были коротенькими. Даже если родственные связи с птерозаврами у него были, шаровиптерикс стал самостоятельной и тупиковой линией развития.

По современным представлениям ближайшим пусть не предком, но родственником предков птерозавров является склеромохл (Scleromochlus taylori). Это была стройная тонкотелая рептилия длиной около 18 см, с длинными лапками. Склеромохл много времени проводил на деревьях, мог передвигаться на двух и четырех ногах, прыгать, и, возможно, имел перепонку, как у белки-летяги.

Отсутствие “переходного звена” усложняет проблему возникновения полета у птерозавров. Возможно, предки птерозавров обитали в заросших лесом ущельях с отвесными стенами. В таких условиях прыжки и парящий полет между ветками и скальными уступами были бы эффективней лазания по ним. Как бы то ни было, к позднему триасу (210 млн л.н.) птерозавры уже поднялись в небо.
Нечисть волосатая
Птерозавры не были первыми летающими животными. Задолго до них летать научились насекомые, достигшие в каменноугольном периоде солидных размеров. Меганевры, стрекозоподобные бестии с размахом крыльев более 70 см, могли бы стать серьезной угрозой, но вымерли за 75 млн лет до появления первых птерозавров.
Этих ранних, триасовых и юрских, летающих рептилий традиционно объединяют в группу рамфоринхоидеи (Rhamphorhynchoidea), противопоставляя поздним, продвинутым птеродактилоидеям (Pterodactyloidea) из мелового периода.

Рамфоринхоидеи в значительной степени сохранили черты строения своих бескрылых предков. Слабая пневматизация скелета, зубастая пасть, длинный хвост, отсутствие гребней на голове – их характерные черты. Размах относительно коротких крыльев был еще невелик, в пределах 2,5 метров. Тело покрывали похожие на волоски пикнофибры, что говорит о теплокровности. «Нечисть волосатая» - так переводится название одного из видов этих животных, Sordes pilosus.
К середине юрского периода рамфоринхоидеи распространяются повсеместно, образуя три экосистемных типа.
К первому типу (видимо, наиболее архаичному) относятся наземно-воздушные хищники, например диморфодон (Dimorphodon). Эти птерозавры довольно много времени проводили на земле, на ветвях деревьев или в скалах. Они хорошо лазали, могли преследовать добычу «пешком», имели широкие короткие крылья, крепкие челюсти и зубы. Их потенциальная добыча – любое мелкое животное, от насекомых до амфибий, ящериц и млекопитающих.

Второй тип представлен охотниками за летающими насекомыми. Могли хватать добычу на лету, были не слишком быстрыми, но весьма маневренными. Из их числа вылетающие по ночам на охоту круглоголовые и большеглазые анурогнатиды (Anurognathidae).



Живые планеры
С середины юрского периода начинается закат рамфоринхоидей, и концу мела сохраняются лишь некоторые реликтовые виды. Их вымирание весьма подозрительно совпадает по времени с появлением и распространением двух новых групп летающих животных.
Первые – потомки самих рамфоринхоидей, птеродактилоидеи. Кости их передних конечностей стали еще более длинными, что позволило увеличить площадь крыла и сократить энергозатраты во время полета. Пропал хвост, вместо него в качестве руля стали использоваться гребни на голове. Достигла максимума пневматизация скелета, что снизило вес животного. В меловом периоде были утрачены зубы, остался только роговой клюв. А еще увеличились доли мозга, отвечающие за движения и координацию. По отношению к своим предками птеродактилоидеи были как истребители середины XX века в сравнении с бипланами Первой Мировой войны.
Ранние птеродактилоиды, такие, как криптодракон (Kryptodrakon progenitor) из формации Шишугоу на северо-западе Китая, имели сопоставимые с рамфоринхоидеями размеры и занимали схожие экологические ниши. Не удивительно, что вскоре они начали теснить своих архаичных собратьев.

Вторая группа – птицы. Оперенные тероподы перешли к полету где-то в лесах Северного Китая 160 – 150 млн лет назад. Поначалу разница в анатомии, образе жизни, способах добычи пропитания позволяла обходиться без прямой конкуренции. Скорее, имело место сдерживающее давление. Более гибкие в эволюционном плане, птицы активно занимали свободные экологические ниши и не пускали туда птерозавров.

Рамфоринхоидеи, зажатые в тиски между птеродактилоидеями и птицами, к началу мелового периода практически исчезли. И уже птеродактилоидеи столкнулись клюв к клюву с птицами. Вытеснить нахальных пернатых не получалось, а вот птицы оказались не прочь при удобном случае занять место рептилий.
И тогда птерозавры начали наращивать собственные размеры. Разница в размерах позволила птеродактилоидеям и птицам занимать одни экосистемы без конкуренции друг с другом. На протяжении всего мелового периода 2 м в размахе крыльев были верхней границей для птиц и нижней для большинства птерозавров.
Выход в крупный размерный класс принес и изменения в экологии.
Тип любителей свежей рыбы и кальмаров сохранился. Но теперь за ними охотились птерозавры с размахом крыльев до 5 – 7 метров. Птеранодоны (Pteranodon), сложив свои длинные крылья, стрелой ныряли в морскую пучину. Никтозавры (Nyctosaurus) с шикарным метровым гребнем на голове выхватывали тонкими зубами рыбу с поверхности воды. Рыбоядные птерозавры были обычными обитателями как морских побережий, так и внутренних водоемов, уживаясь бок о бок с морскими птицами: ихтиорнисами и гесперорнисами.

Претерпел сильные изменения тип наземно-воздушных хищников.
Теперь это были обитатели открытых пространств, ищущие добычу на земле и в водоемах. Одни, как джунгариптериды (Dsungaripteridae), отыскивали в прибрежной полосе моллюсков и ракообразных, дробя их панцири мощными зубами – редкость среди птеродактилоидов. Другие (Istiodactylidae) парили в небе группами, в поисках падали, как это делают стервятники. Были даже фильтраторы наподобие фламинго (Liaodactylus), питающиеся мелкими рачками и планктоном.

А над всеми этими птерозаврами как великаны возвышались представители семейства аждархид. К концу мелового периода они достигли экстремальных для летающих существ размеров. Кетцалькоатль, арамбурджания, хацегоптерикс – крупнейшие представители семейства достигали 10 – 12 м в размахе крыльев, что сравнимо с габаритами самолетов «Цессна». Длинношеие, с огромной головой и острым клювом, они проводили на земле едва ли не больше времени, чем в воздухе. Как гигантские птицы-носороги, вышагивали эти великаны по папоротниковым саваннам. А в случае угрозы расправляли свои огромные кожистые крылья и взмывали в небо.

Итак, дано: не самый крупный кетцалькоатль с размахом крыльев 9,6 м, длиной от носа до кончика задней лапы в 8 м. Стоя на земле это чудище выглядит как крылатый жираф с клювом, а наша цель – заставить его полететь.
Скелет и масса тела
Полет возможен только при оптимальном соотношении массы тела и площади крыла. Оценка массы птерозавров – задача архитрудная: живых аналогов этих существ нет, а экстраполировать показатели птиц и рукокрылых на птерозавров не получается из-за серьезных различий в анатомии.
Для определения массы тела используются трехмерные модели и специальные алгоритмы, высчитывающие объем и ориентировочную плотность различных частей животного. Модель строится на основе палеонтологических реконструкций внешнего вида животного, которые зависят от найденных окаменелостей. Чем более полный скелет можно собрать из его фрагментов, тем более точной будет оценка массы тела.
Современные специалисты по птерозаврам, такие, как Марк Уиттон, дают оценку массы кетцалькоатля в 180 – 250 кг как наиболее реалистичную.
Чтобы иметь размеры с самолет, и весить как кабанчик, птерозаврам пришлось пожертвовать костной тканью.

У птиц масса скелета составляет 8 – 18% от массы тела.
Чем крупнее птица (летающая), тем легче и тоньше ее кости. Такими же тонкими и легкими были кости рамфоринхоидей. У птеродактилоидей они стали совершенно воздушными. И это не метафора. Большинство костей аждархид, включая позвонки, имеют крупные полости. А кости конечностей так и вовсе похожи на полые трубки, толщина стенок которых сравнима с бумажным листом.
По расчетам, скелет нашего кетцалькоатля должен весить всего 18 кг. Из них на передние конечности приходится 8,5 кг, на задние только 1,8 кг. Кости туловища весят 2,7 кг, 2 кг – шейные позвонки, и 3 кг - огромный череп. В общей сложности скелет составляет около 7% массы тела!
У птиц такой недобор костной ткани привел бы к снижению прочности костей крыльев, и их поломке при нагрузках, возникающих во время полета. Но не у птерозавров. Внутри трубчатых костей передних конечностей имелась сложная система перекрещивающихся костных «спиц», придающая всей конструкции огромную прочность. Анализ структуры костной ткани птерозавров говорит о том, что прочностные характеристики превышали показатели птиц в три раза. Это значит, что весь скелет крыла обладал повышенной устойчивостью к нагрузкам.

Крыло и мускулатура
Сами крылья без учета скелета весят примерно 50 кг каждое. Их совокупная площадь порядка 11 м2. Создается впечатление, что крылья недостаточно большие для такого огромного существа. Но тут все дело в нестандартных пропорциях аждархид: слишком большая голова и длинная шея у этих птерозавров.
Крыло птерозавров похоже на крыло рукокрылых, но только внешне. Скелетную основу образуют кости плеча, предплечья, запястья, и чрезвычайно длинный четвертый палец. От запястья отходит птероид, уникальная кость, которая есть только у птерозавров. Его функция – управлять мембраной, расположенную между шеей и запястьем.
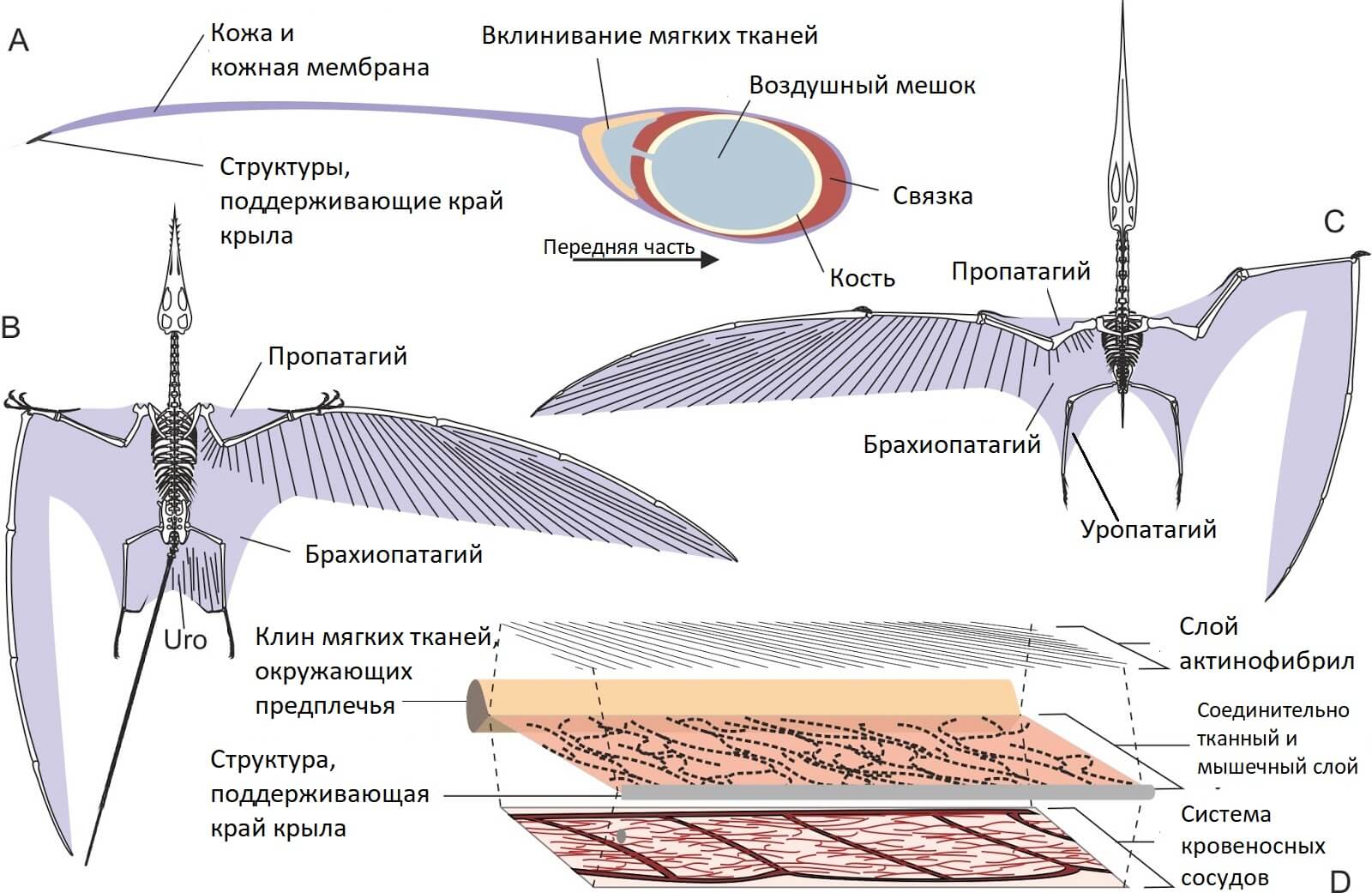
Кожистые мембраны крыльев имеют сложное строение, с прослойкой из соединительной и мышечной ткани и большим количеством кровеносных сосудов. От костной основы к краю крыла проходят особые волокна, актинофибрилы. Пока неизвестен их состав, сложно что-то говорить о назначении этих волокон. Они могут как укреплять крыло, так и менять его геометрию, что влияет на аэродинамические свойства.
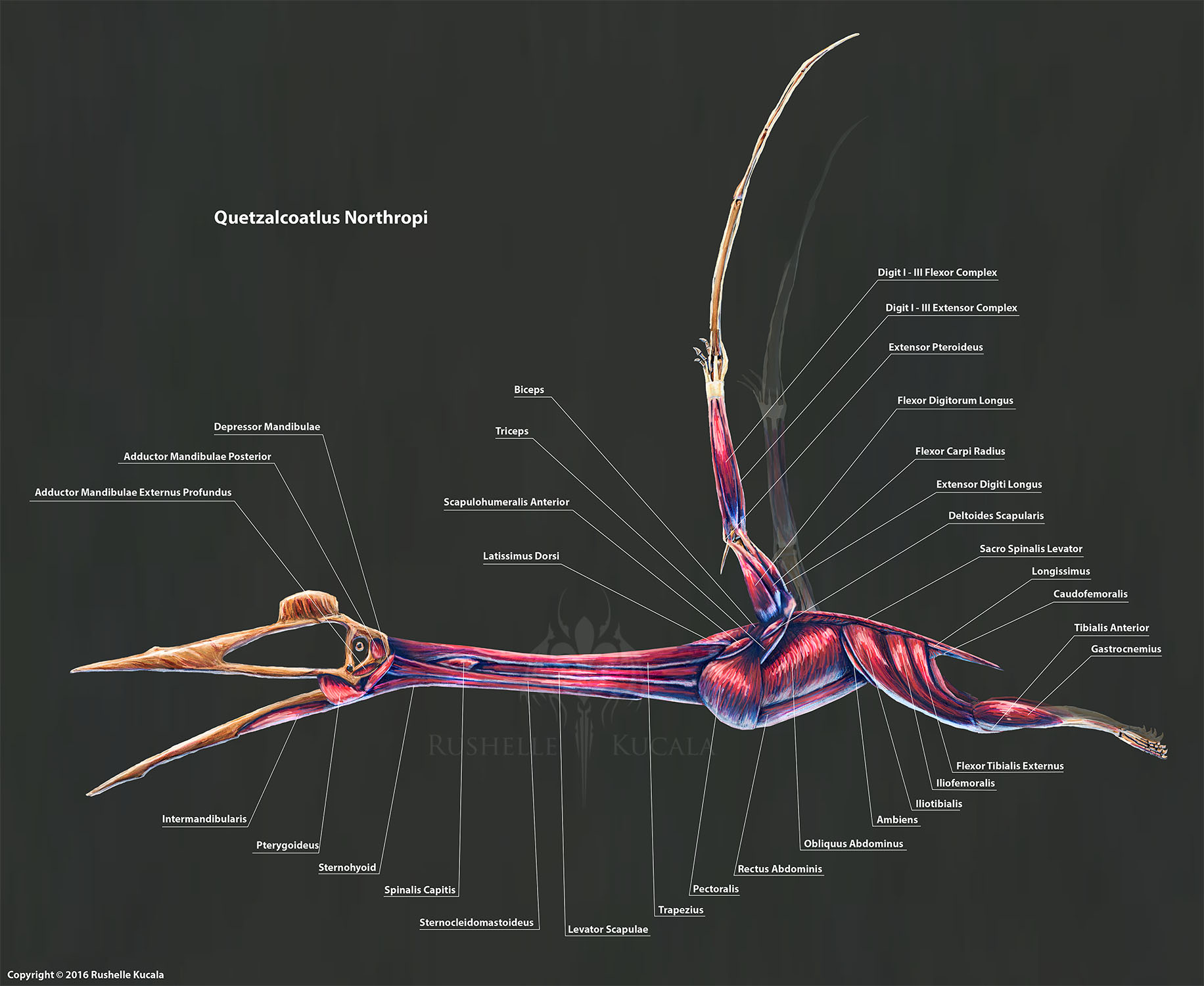
За работу крыльев отвечает мощная мускулатура, вес которой оценивают в 50 кг, что составляет 20% от веса животного. Аналогичные характеристики мы видим у птиц, но распределяется мускулатура верхнего пояса конечностей у кетцалькоатля иначе. За движение крыла отвечают несколько групп мышц, расположенных на груди и спине, и предплечьях, в отличие от компактных грудных мышц птиц. Такое распределение мышечной массы позволяет снизить энергозатраты на машущий полет.
Энергия
Легкие, сердце, пищеварительная система и прочие внутренние органы упакованы в компактное яйцеобразное тело длиной около 1,5 м. Общий вес тела вместе с шеей, головой и задними конечностями около 82 кг.
Активный полет предполагает высокий расход энергии, а следовательно, и быстрый обмен веществ. Сердце – внутренний мотор кетцалькоатля – четырехкамерное, обеспечивающее разделение артериальной и венозной крови и, соответственно, теплокровность. Сложно сказать, с какой частотой билось сердце, перенося к органам и тканям кровь, богатую кислородом. Но о дыхательной системе птерозавров кое что известно.
Позвоночник, ребра и грудина кетцалькоатля образуют достаточно жесткую конструкцию, противостоящую нагрузкам, исходящим от крыльев и их мускулатуры. Сокращение легких в таких условиях обеспечивается не движением ребер, а «печеночным насосом», системой мышц, управляющих движением диафрагмы.
От легких отходят воздушные мешки, которые занимают значительный объем тела, частично проникают в основание крыльев и пронизывают кости кетцалькоатля. Как и у птиц, дыхание осуществляется в два такта: на вдохе воздух заполняет и легкие, и воздушные мешки; на выдохе воздух из воздушных мешков возвращается в легкие. Подобная система обеспечивает высокий уровень кислорода в крови.
И последним элементом энергосистемы кетцалькоатля выступают пикнофибры, покрывающие все его тело. Тепло, выработанное телом, необходимо сохранить. Гибкие, короткие нити, похожие на шерсть выполняют эту задачу.

Управление полетом
Птеродактилоидеи в ходе эволюции утратили хвост, служивший их предкам рулем. Вместо него роль руля на себя взяла крупная голова, украшенная гребнем.
Черепа аждархид – вытянутые, с тонкими беззубыми челюстями, и объединенными носовым и предглазничным отверстием. Область мозга, координирующая сигналы от кожи, мышц, суставов и органов чувств, занимала 7,5% от его массы. Это больше, чем у любого другого животного, включая птиц с их 1-2%.
Полного черепа кетцалькоатля нет, поэтому неизвестны его точная длина или форма гребня. Но исходя из размеров черепов менее крупных аждархид можно сделать вывод, что длина его была не менее 2 метров, а ширина – в пределах 30 см! Узкий, длинный, он выполнял роль руля, помогая птерозавру маневрировать во время полета.
Вес головы с мягкими тканями был явно небольшим, и вряд ли превышал вес черепа более чем в два раза. За управление этой двухметровой плоской «доской» в полете и на земле отвечали мощные шейные мышцы.
Огромная шея длиной 2,5 метра состояла из 9 позвонков, 7 шейных и 2 грудных, утративших ребра. Строение позвонков уникально: позвоночное отверстие смещено к центру, из-за чего позвонок превращается в костную трубку, внутри которой имеется заключенный в кость канал для спинного мозга. Между внешней оболочкой позвонка и мозговым каналом расположены костные пластики, образующие спираль, как спицы в колесе велосипеда, обеспечивающие огромную прочность позвонков.

Летные способности
Для взлета кетцалькоатлю не нужен был трамплин в виде скалы, как это считалось когда-то. Хватало силы ног и крыльев. Птерозавр приседал, сгибая крылья и опираясь на пальцы передних лап, после чего резко отталкивался сильными задними лапами, подпрыгивая вверх. Несколько первых взмахов крыльями были самыми энергозатратными. В целом на активную фазу полета уходило около минуты, в течение которой кетцалькоатль набирал более 500 метров высоты. После чего наступала парящая стадия с использованием восходящих потоков и не требующая большого расхода энергии.
Маневренность полета обеспечивалась движением передних конечностей, меняющим геометрию крыла. Значительную роль в этом играл птероид, кость, отходящая от запястья вперед. Поднимая или опуская его, кетцалькоатль регулировал натяжение передних мембран, что давало возможность быстро менять направление полета. Какую-то роль в аэродинамике играли актинофибрилы, но пока что это тема будущих исследований.

Подводя итог можно сказать, что анатомия, биомеханика, аэродинамика птерозавров – всё было подчинено одной цели, полету.
Возможно ли, что летать могли только молодые особи, а взрослые, с набором веса, проводили всю жизнь на земле? Анализ окаменелостей кетцалькоатлей разных возрастов показывает, что такого не происходило. Даже крупные взрослые животные сохраняли в полной мере характеристики, позволяющие им летать над Землей мелового периода.

